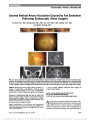
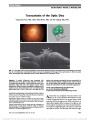
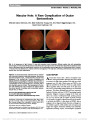
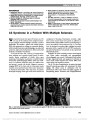
| OCR Text |
Show Williams Fletcher Hoyt William Fletcher Hoyt, MD, received his medical education at the University of Cal-ifornia, San Francisco (UCSF), followed by a year's study at the Wilmer Institute at The Johns Hopkins University, under the mentorship of Frank B. Walsh, MD. Dr. Hoyt re-turned to UCSF in 1958 to found the neuro-ophthalmology service. During a 36-year academic career-all of it at UCSF-he authored 266 journal articles, co-authored (with Frank B. Walsh, MD) the third edition of Clinical Neuro-Ophthalmology, and trained 71 neuro-ophthalmology fellows. In 1983, he received the title of Honorary Doctor of Medicine from the Karolinska Institute. At present, he serves as Professor Emeritus of Ophthalmology, Neurology, and Neurosurgery at UCSF. In recognition of his con-tributions, in 2001 the North American Neuro-Ophthalmology Society, in conjunction with the American Academy of Ophthalmology, initiated the Hoyt Lecture to be delivered each year at the Annual Meeting of the American Academy of Ophthalmology. Are We There Yet? Is Neuro-Ophthalmology at the Cusp of a Paradigm Shift? Lessons From Leber Hereditary Optic Neuropathy Alfredo A. Sadun, MD, PhD Journal of Neuro-Ophthalmology 2013;33:189-197 doi: 10.1097/WNO.0b013e31829343a0 © 2013 by North American Neuro-Ophthalmology Society It is a great honor and pleasure and very humbling to deliver this lecture. I stand in awe of William Hoyt and the august list of past awardees. Most of my training in neuro-ophthalmology was with Simons Lessell and my debt to him can't be overstated. One of the many kindnesses he did for me was to send me to San Francisco to spend the last 6 weeks of my fellowship with Dr. Hoyt (see Supplemental Digital Content, Figure E1, http://links.lww.com/WNO/A78). I am also humbled by the prospect of discussing the topic I may have foolishly chosen for myself. Are we there yet? Is neuro-ophthalmology at the cusp of a paradigm shift? This comment is in reference to the classical work of Thomas Kuhn. In 1962, Kuhn, a physicist, a philosopher, and a professor at Massachusetts Institute of Technology published "The Structure of Scientific Revolutions" (1). In this book, he described what became popularly known as the "paradigm shift." Kuhn pointed out that scientists accumulate evidence that supports their model of the real world. However, such evidence eventually becomes messy and even self-contradictory until at such time as the revolution, or paradigm shift, re-establishes a fundamentally different model. Kuhn described 3 stages for each science as: the central paradigm, the anom-alous elements, and the reconfiguration of the new paradigm. What has all this got to do with neuro-ophthalmology? Well, the central paradigm for neuro-ophthalmology was that a single physical lesion produced a classical set of signs and symptoms. However, with the arrival of neuroimaging, it appeared that computed tomography and magnetic reso-nance imaging might do the job better than a neuro-ophthalmologist. In fact, when I was with Dr. Hoyt, I recall him bemoaning the possible end of neuro-ophthalmology with the success of neuroimaging. However, there are diseases that do not conform to the localizing lesion paradigm and are not detectable with neuroimaging techniques. Departments of Ophthalmology and Neurosurgery, Doheny Eye Institute, Keck School of Medicine of University of Southern Cal-ifornia, Los Angeles, California. Supported by the International Foundation of Optic Nerve Diseases, Struggling with Lebers, the Poincenot Family for LHON, the Eierman Foundation, Research to Prevent Blindness, and National Institutes of Health Grant EY03040. The author reports no conflicts of interest. The author has consulted for Pfizer Pharmaceuticals and has con-ducted Leber hereditary optic neuropathy-related research with an unrestricted grant from Edison Pharmaceuticals. Supplemental digital content is available for this article. Direct URL citations appear in the printed text and are provided in the full text and PDF versions of this article on the journal's Web site (www. jneuro-ophthalmology.com). Address correspondence to Alfredo A. Sadun, MD, PhD, Doheny Eye Institute, Keck School of Medicine of USC, 1450 San Pablo Street, Los Angeles, CA 90033; E-mail: asadun@usc.edu Sadun: J Neuro-Ophthalmol 2013; 33: 189-197 189 The Eleventh Annual Hoyt Lecture Copyright © North American Neuro-Ophthalmology Society. Unauthorized reproduction of this article is prohibited. MITOCHONDRIAL OPTIC NEUROPATHIES The metabolic injury that characterizes mitochondrial optic neuropathies (MON) might be characterized as a biochem-ical paradigm. MON can be genetic, as in Leber hereditary optic neuropathy (LHON) or dominant optic atrophy, secondary to nutritional deficiencies, such as with vitamin B12 or folic acid, or due to a variety of toxic agents, many of which are antibiotics that interfere with both bacterial and mitochondrial ribosomes (2,3). The prototypic MON, LHON, was first described in 1858 by Von Graefe. In 1871, Leber reported 4 families and proposed that this was a hereditary and probably X-linked disease (4-7). However, it wasn't until 1988 when Wallace et al (8) described the 11778 mitochondrial DNA (mtDNA) mutation that we began to get a handle on both the genetics and the pathophysiology of this MON. Neuro-ophthalmologists are well aware of the typical pre-sentation of LHON. But youmight find it interesting to know that what we often see in the United States is a little different in Brazil. In the United States, I might see a young man in good health without a family history of blindness. He goes off to college and adopts a lifestyle, which includes smoking and heavy drinking. A few months later he suffers subacute loss of vision in one eye and is seen at the student health center and diagnosed with optic neuritis. The work-up for multiple sclerosis is entirely negative and within a month or two he loses vision in the felloweye. It's about then that he gets referred to neuro-ophthalmology and the diagnosis becomes clear. I'll tell you later how this plays out a little bit differently in Brazil. FOUR FUNDAMENTAL QUESTIONS There are many mysteries in LHON. Let me point out 4: 1. Why are some individuals with the mutation affected and others not? (The problem of penetrance.) 2. Why are men much more likely to lose vision than women? (The problem of gender bias.) 3. Why is it almost always just the optic nerve that suc-cumbs? (The problem of tissue specificity.) 4. Why does the catastrophic loss of vision occur during young adulthood? (The problem of mass tipping point.) Mitochondria are remarkable organelles. We tend to describe them as individual little beans that float freely in the cytoplasm but of course that isn't really accurate. Mito-chondria usually fuse into a syncytium, which follows the cytoskeleton and then sometimes they undergo fission, per-haps for special functions such as transport. The mitochondria coordinate with the nucleus in a variety of functions, includ-ing their own reproduction, which occurs in the soma of the retinal ganglion cell (RGC) in proximity to the nucleus. They may then hitch a ride on kinesin and dynein motors, to travel down the axons and provide adenosine triphosphate (ATP) to distal elements. In producing ATP, mitochondria utilize oxi-dative phosphorylation and, as a consequence, produce great amounts of reactive oxygen species (ROS) (9,10). LHON PEDIGREE IN BRAZIL I was extremely fortunate to lead a team of international investigators, numbering up to 40 at a time, in yearly field investigations to rural Brazil. We utilized an ophthalmology clinic in Colatina, Brazil, and received support from many volunteers organized through the Federal University of Sao Paulo (11). The first such visit in 2001 allowed us to define a large pedigree of 11778 LHON. In subsequent years, we worked out the psychophysics, came to understand that there was subclinical disease in carriers of LHON, monitored the natural history of conversions, and established a number of psychophysical, clinical, subclinical, and serum biomarkers (12-18). We characterized a pedigree with 362 members from 8 generations that all began in 1861 in Verona, Italy (Fig. 1). From there, a 16-year-old girl immigrated to Brazil to become the founder of this family. Almost all her progeny have remained in the same area in rural Brazil (11). All of this came to our attention through a series of e-mails that originated from the mother of a 14-year-old boy who had just lost vision in one eye. This mother recalled that 30 years before, her 2 older brothers had experienced similar profound visual loss. By way of the Internet, she resolved to better understand her family's disease and then to contact experts in the field whom she hoped could provide a means of keeping her child from going blind in the second eye. Through the International Foundation for Optic Nerve Diseases, she found me and, despite the complications of language and culture, we FIG. 1. Leber hereditary optic neuropathy pedigree com-posed of 8 generations. The affected individual in generation 2 was born in 1861 and emigrated from Verona to the state of Spirito Santo, Brazil, at the age of 16 years, where she became the founder of the seventh-generation 332-member pedigree. Another 30 members still living in Italy have also been identified (adapted from Sadun et al (14)). 190 Sadun: J Neuro-Ophthalmol 2013; 33: 189-197 The Eleventh Annual Hoyt Lecture Copyright © North American Neuro-Ophthalmology Society. Unauthorized reproduction of this article is prohibited. concluded that the diagnosis that she proposed, LHON, was correct. She told of a 100 family members who might be carriers of LHON with several having already lost vision, and she was anxious that we evaluate her son. We resolved to assemble a large team to conduct a field investigation in Brazil.We arranged for her, with her son and 2 brothers, to be evaluated at the Federal University in Sao Paulo. Measure-ments of visual acuity, visual fields, and fundus photographs were sent to us along with blood for genetic testing. Unfor-tunately, by the time that they arrived in Sao Paulo, her 14-year-old son had already lost vision in the second eye. He had visual acuity of 20/200, right eye, and 20/800, left eye, and his fundus appearance is shown in Figure 2. The logistics were challenging to bring expertise and sophisticated equipment from around the world into a remote area of Brazil. We managed these for 11 consecutive years and made a number of interesting findings. For example, although we found that the average age of onset for the entire family was 29 years, subgroup analysis showed this age to be 23 years for men and 33 years for women. Women had a bimodal distribution, with some losing vision at about the same age as the men but others losing vision as they approached menopause. The visual acuity of those who became affected and lost vision averaged 5/400. Remarkably, the penetrance rate was much higher if the mother herself had lost vision. In other words, if a mother lost vision, it was almost certain that her sons would be affected and her daughters were at high risk. But if a mother was an unaffected carrier, her daughters were very unlikely to lose vision and even her sons had good odds of preserving their sight. Similar to reports from other pedigrees (19-24), we found a male to female ratio of about 4:1 in this 11778 family. It was intriguing that the most common month for vision loss in Brazil was April. We will later revisit this FIG. 2. Leber hereditary optic neuropathy index case of a 14-year-old boy. There is pseudooptic disc edema and retinal nerve fiber swelling (arrows) in the both eyes (left and right), mild left optic disc pallor, and decreased nerve fiber layer in the papillomacular bundle of each eye (adapted from Sadun et al (12)). FIG. 3. Light (A) and electron microscopic (B) photographs of fibers in the retrobulbar optic nerve from a 76-year-old woman with Leber hereditary optic neuropathy who had lost vision about 50 years ago. A. The remaining fibers (arrows) are swollen and beaded. B. Some fibers are actively undergoing degeneration (arrow) (A, paraphenylene diamine stain; scale bar = 10 mm; B, scale bar = 1 mm). Sadun: J Neuro-Ophthalmol 2013; 33: 189-197 191 The Eleventh Annual Hoyt Lecture Copyright © North American Neuro-Ophthalmology Society. Unauthorized reproduction of this article is prohibited. seasonal phenomenon in the light of the likely 6-week lag period between the triggering insult and the loss of vision in LHON. We found that the likelihood of conversion from carrier status to affected was much higher in those exposed to pesticides, cigarette smoking, and alcohol (11,12). Sixty-five percent of our affected LHON patients smoked compared with 26.1% of the off-pedigree controls and 13.5% of our LHON carriers. With regards to the consumption of at least 2 drinks of alcohol per day, LHON affected (60%) were much more likely to consume compared with controls (38.2%) or carriers (33.8%). Thus, there was a statistically significant increased rate of smoking and alcohol use in LHON affected compared with either carriers or controls. After examining close to 300 members of the LHON pedigree every year for 11 years, what did we learn? Our conclusions can be summarized as follows: Although there is an apoplectic loss of vision that occurs over a period of weeks to months, there is also clinical evidence of further progression that continues long term, year after year. This progression is hard to monitor because visual acuity does not continue to change but there is progressive visual field loss well beyond the central 30° angle. One lesson here is that you should keep seeing these patients, keep getting visual fields, and try to use visual field strategies that extend beyond 30° angle. This late progression is probably life-long. We obtained histopatholog-ical tissue from a 76-year-old patient who had lost her vision about 5 decades earlier. Her vision had been estimated as counting fingers for decades before she died of cardiovascular complications. In examination of her optic nerves, there was still active degeneration of some remaining fibers found in the superonasal quadrant, shown by light and electron micros-copy (EM) (Fig. 3). The appearance on EM reflects degen-eration that occurred in the last 4-6 days of her life or 50 years after she first noted visual loss! What about the asymptomatic carriers? These patients may be asymptomatic but they often harbor subclinical disease. We found zones of mild sectoral disc edema, sometimes with telangiectatic vessels, in more than one-third of the carrier cases (Fig. 4) (15). Using more sophisticated psychophysical techniques (Cambridge color system), we were able to dem-onstrate some dyschromatopsia in about half of the carriers (16). This dyschromatopsia may be too subtle to show up on standard Ishihara color test plates. Finally, optical coherence tomography (OCT) turned out to be an excellent means of discerning structural changes in asymptomatic carriers. Several female carriers and almost 90% of male carriers demonstrated retinal nerve fiber layer (RNFL) thickening in the inferotem-poral quadrant (25-27). In this extensive longitudinal study, OCT was the most reliable method to demonstrate structural changes, which preceded and then reached a crescendo at the time of visual loss. This began with swelling of the peripapillary RNFL in the inferotemporal quadrant and was followed by swelling superiorly and temporally. Eventually, each quadrant took turns in first swelling and then undergoing atrophy. Barboni et al (26,27) demonstrated this RNFL thickening in carriers and the orderly wave of thickening followed by atrophy that occurs during conversion to affected status. However, not all LHON carriers with OCT changes or pseudo-disc edema lost vision. Some waxed and waned for many years and some even improved without any significant visual loss. REACTIVE OXYGEN SPECIES Many LHON patients in Brazil experienced vision loss each year in April. In our discussions, it became evident that the visual loss was preceded, in February, by attendance at Carnival, a week-long festivity in Brazil characterized by FIG. 4. Twelve-year-old male carrier with 11778 Leber hereditary optic neuropathy. The patient had no visual com-plaints with visual acuity of 20/20 bilaterally. Yet there is retinal nerve fiber layer swelling associated with telangiec-tatic vessels (arrows). This patient underwent subacute loss of vision about 2 years after this photograph was taken. FIG. 5. Sagittal sections (lamina cribrosa at the top) of plastic embedded normal human optic nerve (A) and from a patient with Leber hereditary optic neuropathy (LHON) (B). Optic nerve staining with nitrotyrosine immunolabeling is light in normal optic nerve and heavy (arrowheads) in LHON patient (scale bars = 200 mm). 192 Sadun: J Neuro-Ophthalmol 2013; 33: 189-197 The Eleventh Annual Hoyt Lecture Copyright © North American Neuro-Ophthalmology Society. Unauthorized reproduction of this article is prohibited. a great deal of binge drinking. In most cases, the carrier was a teenager who, for the first time, was allowed to accompany a family member to Carnival where they both participated in heavy consumption of alcohol. The Brazil longitudinal studies take into consideration a number of epidemiological findings that suggest both genetic and environmental risk factors for conversion. Gene linkage analysis points to a "hot spot" on the X chromosome, which may harbor a nuclear modifying factor. There are many genes in this area. One candidate gene is for manganese superoxide dismutase (MnSOD) or SOD2, a specific form of superoxide dismutase used by mitochondria to help neutralize ROS. Of course, ROS may also come from the environment introduced by activities such as heavy drinking and exposure to smoke. We reported several members of the same LHON family who lost vision while being exposed to smoke from a tire fire (28). This vulnerability to ROS has also been modeled in cybrid experiments. Cybrids are cell cultures that take advantage of an immortalized cell line, such as osteo-sarcoma, in which the original mitochondria have been destroyed and replaced with mitochondria obtained from the white blood cells taken from LHON patients. These cultures can be grown indefinitely and harbor only the mitochondria of interest. With these experiments, it has been shown that the 3 primary LHON mutations only produce a modest (approximately 20%) decrease in oxygen consumption and ATP production but proportionately a much greater increase in ROS production (29-32). To look further at the role of ROS, we looked for the presence of nitrotyrosine in optic nerves from LHON patients. Nitrotyrosine is a product of accumulated ROS damage. As demonstrated in our optic nerve sections, nitrotyrosine immunostaining is minimal in control optic nerves and extreme in nerves from patients with LHON (Fig. 5). This suggests a life-long exposure to high levels of ROS in LHON. ROS overproduction makes sense regarding the patho-physiology that leads to RGC death in LHON, for we know that the mtDNA mutation leads to insufficiency at complex I. This produces a small decrease in ATP but a very large increase in ROS (29,32). Smoke or drinking of alcohol can add significant amounts of ROS, which may then exceed a certain threshold, causing apoptotic cell death (33,34). Furthermore, RGCs may be at particular risk because of their long unmyelinated intraretinal segment. Mitochondria are likely the lynch pin of this pathological process that ends with RGC apoptosis. To gain greater insight into the nature of mitochondria, let us consider their origin. The well-known theory of Margulis brings us back billions of years ago to when the world was populated by aerobic bacteria, anaerobic eukaryotes, and blue green algae. The blue green algae slowly brought the oxygen levels in the atmosphere from as low as 2% to about 22% (35). This was catastrophic for the anaerobic eukaryotes, which could not easily survive these high oxygen levels. Fortuitously, as Margulis et al (36) suggested, a small miracle of life occurred, which she termed "endosymbiosis." That is to say, a eukaryote consumed but did not destroy a bacterium that it had in-gested, and this surviving bacterium went on to become a pro-to mitochondrion. This unified cell prospered with the mitochondria providing relief from the oxygen and an FIG. 6. Three potential triggers of mitochondrial optic neuropathy lead to both a reduction of adenosine triphosphate (ATP) and accumulation of reactive oxygen species (ROS) that can, by changing the mitochondrial membrane's electrical potential, open the mitochondrial permeability transition pore (MPTP) and trigger apoptosis (modified from Sadun and Wang (2)). Sadun: J Neuro-Ophthalmol 2013; 33: 189-197 193 The Eleventh Annual Hoyt Lecture Copyright © North American Neuro-Ophthalmology Society. Unauthorized reproduction of this article is prohibited. efficient source of ATP for the new amalgam eukaryote. Mito-chondria represent vestige bacteria within the modern eukary-ote. The eukaryote, with many sophisticated subsystems and organelles, brought a lot to the table as well. Specifically, the modern eukaryote provided a safe haven in the form of a nucleus that was sheltered away from the high ROS levels naturally generated by mitochondria. With this cooperative system, some eukaryotes eventually became the multicellular organisms, including humans, that now populate the earth. This leads us to ask a few more questions about LHON. Why does blocking oxidative phosphorylation in mitochon-dria lead to apoptosis? Why the tissue specificity (optic nerve) and why the predilection for the fibers of the papillomacular bundle (PMB)? Once again we have to think of the Margulis paradigm and understand that for a billion years preceding this symbiosis, bacteria were constantly destroying and being destroyed by eukaryotes (35). Consequently, each type of cell developed an arsenal of biochemical weapons as part of an arms race with the other. After the symbiosis, the DNA library for this arsenal sat as an unused time bomb. But this time bomb could be set off by unleashing the machinery of the protomitochon-dria. Generally, there is not much selection advantage for a single cell to commit suicide. However, once multicellu-larity occurred, this system became an extremely useful tool as part of what we now term apoptosis; it permitted a variety of key processes such as ontogeny, which allowed for the development of organs and appendages, and immunosur-veillance for the consequent destruction of cancer cells. Developmental neuronal specificity can occur by the selec-tive pruning of neurons based on their connections, which, if appropriate, provide chemical signals that modulate apo-ptosis. RGCs use this method as they go from 5 million to about 1 million during fetal development (37). How does the mitochondrion control one aspect of apoptosis? The inner membrane of the mitochondrion has an electrical potential. This electrical potential is altered by a variety of signals. ROS lower the electrical potential of the membrane, which, at a key voltage gate, can lead to opening of the mitochondrial permeability transition pore (MPTP) (33,34). Once the MPTP opens, it releases cytochrome c, which in turn permits pro-caspase 9 to become caspase 9, and the entire cascade of apoptosis can be triggered (Fig. 6). WHAT MAKES THE RGCs VULNERABLE? The brain weighs approximately 2% of the body weight and yet consumes 20% of its oxygen (38). This reflects the extremely high energy needs required to maintain neurons and, specifically, axons, which bear the high-energy costs FIG. 7. Six optic nerves from 1 normal and 5 Leber hereditary optic neuropathy (LHON) patients have been oriented such that the temporal side is to the left. They have been graded from normal (0-upper left) to mild (1) to grades of moderate to very severe (5-lower right) optic atrophy. The brown staining shows the fascicles of myelinated axons in epon embedded sections. There is a specific pattern of increasing atrophy extending from the papillomacular bundle to involve the entire optic nerve with only sparing of axons in the supranasal quadrant (paraphenylenediamine stain; scale bars = 500 mm). The number in the equation of mitochondrial stress index shows the number of nodes of Ranvier (N#) times the length of the node (NL) times the circumference of the axon (NC) at the node. The denominator is the number of mitochondria (mt#) that exist to sustain this surface area. The equation can be reduced assuming that the retinal nerve fiber layer acts as a giant node of Ranvier. Substituting for circumference and area the equation reduces further to 2/r (r = radius of the axon). The smallest fibers are predicted to be involved first in a mitochondrial optic neuropathy and this holds true in LHON. 194 Sadun: J Neuro-Ophthalmol 2013; 33: 189-197 The Eleventh Annual Hoyt Lecture Copyright © North American Neuro-Ophthalmology Society. Unauthorized reproduction of this article is prohibited. of membrane repolarization following every action poten-tial. This is minimized by saltatory conduction made pos-sible by myelination, which not only allows for faster conduction velocity but also a greater economy of energy expenditure and hence less strain on the greatest source of ATP-the mitochondria. However, this is problematic for RGCs whose axons, that must be transparent, run an unmyelinated course in the retina. We know from the usual clinical presentation of MON (poor visual acuity, dyschromatopsia, and central scotomas) that the PMB is selectively and initially affected (7). In LHON, not only is there the tissue specificity of the optic nerve but more selective still, early involvement of the PMB. We hypothe-sized that this is due to the small size of the fibers that constitute the PMB. We calculated that the energy required for repolarization of an axon potential should be the num-ber of nodes (of Ranvier) times the average length of each node times the axonal circumference at the node (Fig. 7). This represents the numerator of an equation reflecting the mitochondrial axonal stress, where the denominator would be the number of mitochondria that can provide these quantities of ATP. This latter number is limited by the volume of the axon cylinder, which is calculated as the length times pr2, where r is the radius of the axon (small in the PMB). Further reduction of this equation leads to an index that goes as 2/r. The question then is, can we predict the areas of optic nerve involvement based solely on the distribution of axon fiber size? To test this hypothesis, approximately 100,000 axons from normal human optic nerves were measured in 32 regions. The fibers in each region and then the regions were ranked according to size (producing a series of spectra). This was painstaking work by Pan et al (39) and included exten-sive computations. The histograms of the nerve fiber spectra showed a narrow range of small fibers coming from the PMB that were found in the inferotemporal regions (Fig. 8A) and a broader range of larger fibers coming from the superonasal regions (Fig. 8B). Seen another way, a gray scale representation of a normal human optic nerve shows that the smallest fibers are inferotemporal and central, and larger fibers are peripheral, and especially superonasal; this matches exactly the distribution as seen in the patterns of degeneration in LHON optic nerves (Fig. 7). In Figure 9, we have displayed a schematic superimposing on an optic nerve from a LHON patient with only mild pathology, the rank order distribution of fibers by size. The smallest fibers are found in the areas numbered 1 and the largest in the areas numbered 5. These numbers 1-5 also represent the order of progression of LHON degeneration and as pre-dicted by the mitochondrial stress index equation. FIG. 8. Control human optic nerve axons cut in cross section. Arrows lead to the respective region from which the photo-micrograph was captured. A. The image reveals relative uniformity of small axonal size within the inferotemporal optic nerve. B. The superonasal optic nerve shows a much more heterogenous pattern of axonal diameters with a tendency toward larger axons (paraphenylenediamine stain; scale bar = 5 mm). Schematic representation of a control human optic nerve cut in cross section showing nerve fiber layer spectrum histograms from each region, based on 100,000 fibers captured and measured (39). In the histograms, the x axis is radius of each axon and y axis is number of fibers. With greater number of large axons on the side of the optic nerve, the spectra are shifted to the right. Sadun: J Neuro-Ophthalmol 2013; 33: 189-197 195 The Eleventh Annual Hoyt Lecture Copyright © North American Neuro-Ophthalmology Society. Unauthorized reproduction of this article is prohibited. CONCLUDING THOUGHTS So, are we there yet? Well, research in LHON and other MONs has moved in several important fronts leading to the consideration of new treatment options such as quinones that might potentially redirect the electron that spills off complex I back into complex III and reduce some ROS production. Recent publications have described the use of a second-generation quinone, idebenone, with some success (40-43). A third-generation quinone, EPI-743, has shown that, at least in some cases, there may be significant amelioration of vision and visual field loss in cases of LHON (44,45). But are we there yet? I would have to say not quite, insofar as we lack testing on an appropriate animal model. But we have good reason to think that we are close. We and other groups are conducting such research on recently developed faithful animal models of LHON (46,47). To paraphrase Enrico Fermi, who was describing the great advances in physics in the 20th century, "theory stim-ulates experimentation which in turn stimulates new theory." And so yesterday's discovery becomes today's tool. Although neuro-ophthalmology is not physics, and our paradigm shift might not be as dramatic, we do well in avoiding polemics. We will watch with delight as the next generation of inves-tigators take what we've done to a whole new level. ACKNOWLEDGMENTS Much of this work has been in close collaborations with the author's colleagues. The author thanks Fred Ross-Cisneros, lab supervisor, who has patiently pushed himself to the limit in generating critical data and the perfect photomicrographs. The author thanks Michelle Wang for meeting every deadline and Billy Pan who spent countless hours measuring hundreds of thousands of axons. The author particularly thanks scientific siblings, Valerio Carelli and Chiara La Morgia, and the core group of field investigators of the Brazil-LHON team, Solange Salomao, Adriana Berezovsky, Rubens Belfort, Milton Moraes (father and son), Dora Ventura, Peter Quiros, Piero Barboni, Filipe Chicani, Federico Sadun, and Anna Maria DeNegri. The author also thanks the others who participated in the field investigation team. Finally, the author thanks the hun-dreds of members of the Brazilian family who are descendants of that intrepid immigrant, Maria Franchi. REFERENCES 1. Kuhn TS. The Structure of Scientific Revolutions. Chicago, IL: University of Chicago Press, 1962. 2. Sadun AA, Wang MY. Ethambutol optic neuropathy: how we can prevent 100,000 new cases of blindness each year. J Neuroophthalmol. 2008;28:265-268. 3. Sadun AA, Wang MY. Drug related mitochondrial optic neuropathies. J Neuroophthalmol. 2013;33:172-178. 4. Carelli V, Ross-Cisneros FN, Sadun AA. Optic nerve degeneration and mitochondrial dysfunction: genetic and acquired optic neuropathies. Neurochem Int. 2002;40:573-584. 5. Leber T. Uber hereditare und congenitalangelegte sehnervenleiden. Albrecht von Graefes Archiv fuer Ophthalmol. 1871;17:249-291. 6. Biousse V, Newman NJ. Neuro-ophthalmology of mitochondrial diseases. Curr Opin Neurol. 2003;16:35-43. 7. Carelli V, Ross-Cisneros FN, Sadun AA. Mitochondrial dysfunction as a cause of optic neuropathies. Prog Retin Eye Res. 2004;23:53-89. 8. Wallace DC, Singh G, Lott MT, Hodge JA, Schurr TG, Lezza AM, Elsas LJ 2nd, Nikoskelainen EK. Mitochondrial DNA mutation associated with Leber's hereditary optic neuropathy. Science. 1988;242:1427-1430. 9. Sena LA, Li S, Jairaman A, Prakriya M, Ezponda T, Hildeman DA, Wang CR, Schumacker PT, Licht JD, Perlman H, Bryce PJ, Chandel NS. Mitochondria are required for antigen-specific T cell activation through reactive oxygen species signaling. Immunity. 2013;14:S1074-S7613.(13)00051-4. 10. Knockaert L, Descatoire V, Vadrot N, Fromenty B, Robin MA. Mitochondrial CYP2E1 is sufficient to mediate oxidative stress and cytotoxicity induced by ethanol and acetaminophen. Toxicol In Vitro. 2011;25:475-484. 11. Sadun AA, Carelli V, Salomao SR, Berezovsky A, Quiros PA, Sadun F, DeNegri AM, Andrade R, Moraes M, Passos A, Kjaer P, Pereira J, Valentino ML, Schein S, Belfort R. Extensive investigation of a large Brazilian pedigree of 11778/ haplogroup J Leber hereditary optic neuropathy. Am J Ophthalmol. 2003;136:231-238. 12. Sadun AA, Carelli V, Salomao SR, Berezovsky A, Quiros P, Sadun F, DeNegri AM, Andrade R, Schein S, Belfort R. A very large Brazilian FIG. 9. Cross section of optic nerve from a mild case of Leber hereditary optic neuropathy. Superimposed are num-bers (1-5) that represent the fiber calibers ranked by size. These numbers (1-5) also represent the stages of involve-ment/ severity as a wavefront that begins on the temporal side (papillomacular bundle), then, in stages, progresses to the nasal side until only the superonasal region is relatively spared. Compare with cross-sectional photographs in Figure 7 (paraphenylenediamine stain; scale bar = 500 mm). 196 Sadun: J Neuro-Ophthalmol 2013; 33: 189-197 The Eleventh Annual Hoyt Lecture Copyright © North American Neuro-Ophthalmology Society. Unauthorized reproduction of this article is prohibited. pedigree with 11778 Leber's hereditary optic neuropathy. Trans Am Ophthalmol Soc. 2002;100:169-178; discussion 178-179. 13. Sacai PY, Salomão SR, Carelli V, Pereira JM, Belfort R Jr, Sadun AA, Berezovsky A. Visual evoked potentials findings in non-affected subjects from a large Brazilian pedigree of 11778 Leber's hereditary optic neuropathy. Doc Ophthalmol. 2010;121:147-154. 14. Sadun AA, Salomao SR, Berezovsky A, Sadun F, Denegri AM, Quiros PA, Chicani F, Ventura D, Barboni P, Sherman J, Sutter E, Belfort R Jr, Carelli V. Subclinical carriers and conversions in Leber hereditary optic neuropathy: a prospective psychophysical study. Trans Am Ophthalmol Soc. 2006;104:51-61. 15. Sadun F, De Negri AM, Carelli V, Salomao SR, Berezovsky A, Andrade R, Moraes M, Passos A, Belfort R, da Rosa AB, Quiros P, Sadun AA. Ophthalmologic findings in a large pedigree of 11778/Haplogroup J Leber hereditary optic neuropathy. Am J Ophthalmol. 2004;137:271-277. Erratum in: Am J Ophthalmol. 2004; 137: following 793. 16. Ventura DF, Quiros P, Carelli V, Salomão SR, Gualtieri M, Oliveira AG, Costa MF, Berezovsky A, Sadun F, de Negri AM, Sadun AA. Chromatic and luminance contrast sensitivities in asymptomatic carriers from a large Brazilian pedigree of 11778 Leber hereditary optic neuropathy. Invest Ophthalmol Vis Sci. 2005;46:4809-4814. 17. Ventura DF, Gualtieri M, Oliveira AG, Costa MF, Quiros P, Sadun F, de Negri AM, Salomão SR, Berezovsky A, Sherman J, Sadun AA, Carelli V. Male prevalence of acquired color vision defects in asymptomatic carriers of Leber's hereditary optic neuropathy. Invest Ophthalmol Vis Sci. 2007;48:2362-2370. 18. Yee KM, Ross-Cisneros FN, Lee JG, Da Rosa AB, Salomao SR, Berezovsky A, Belfort R Jr, Chicani F, Moraes-Filho M, Sebag J, Carelli V, Sadun AA. Neuron-specific enolase is elevated in asymptomatic carriers of Leber's hereditary optic neuropathy. Invest Ophthalmol Vis Sci. 2012;53:6389-6392. 19. Newman NJ. From genotype to phenotype in Leber hereditary optic neuropathy: still more questions than answers. J Neuroophthalmol. 2002;22:257-261. 20. Yu-Wai-Man P, Griffiths PG, Chinnery PF. Mitochondrial optic neuropathies-disease mechanisms and therapeutic strategies. Prog Retin Eye Res. 2011;30:81-114. 21. Newman NJ. Hereditary optic neuropathies: from the mitochondria to the optic nerve. Am J Ophthalmol. 2005;140:517-523. 22. Fraser JA, Biousse V, Newman NJ. The neuro-ophthalmology of mitochondrial disease. Surv Ophthalmol. 2010;55:299-334. 23. Nikoskelainen EK, Huoponen K, Juvonen V, Lamminen T, Nummelin K, Savontaus ML. Ophthalmologic findings in Leber hereditary optic neuropathy, with special reference to mtDNA mutations. Ophthalmology. 1996;103:504-514. 24. Spruijt L, Kolbach DN, de Coo RF, Plomp AS, Bauer NJ, Smeets HJ, de Die-Smulders CE. Influence of mutation type on clinical expression of Leber hereditary optic neuropathy. Am J Ophthalmol. 2006;141:676-682. 25. Ramos Cdo V, Bellusci C, Savini G, Carbonelli M, Berezovsky A, Tamaki C, Cinoto R, Sacai PY, Moraes-Filho MN, Miura HM, Valentino ML, Iommarini L, De Negri AM, Sadun F, Cortelli P, Montagna P, Salomao SR, Sadun AA, Carelli V, Barboni P. Association of optic disc size with development and prognosis of Leber's hereditary optic neuropathy. Invest Ophthalmol Vis Sci. 2009;50:1666-1674. 26. Barboni P, Carbonelli M, Savini G, Ramos Cdo V, Carta A, Berezovsky A, Salomao SR, Carelli V, Sadun AA. Natural history of Leber's hereditary optic neuropathy: longitudinal analysis of the retinal nerve fiber layer by optical coherence tomography. Ophthalmology. 2010;117:623-627. 27. Barboni P, Savini G, Feuer WJ, Budenz DL, Carbonelli M, Chicani F, Ramos CD, Salomao SR, De Negri A, Parisi V, Carelli V, Sadun AA. Retinal nerve fiber layer thickness variability in Leber hereditary optic neuropathy carriers. Eur J Ophthalmol. 2012;4:985-991. 28. Sanchez RN, Smith AJ, Carelli V, Sadun AA, Keltner JL. Leber hereditary optic neuropathy possibly triggered by exposure to tire fire. J Neuroophthalmol. 2006;26:268-272. 29. Carelli V, Vergani L, Bernazzi B, Zampieron C, Bucchi L, Valentino M, Rengo C, Torroni A, Martinuzzi A. Respiratory function in cybrid cell lines carrying European mtDNA haplogroups: implications for Leber's hereditary optic neuropathy. Biochim Biophys Acta. 2002;1588:7-14. 30. Ghelli A, Zanna C, Porcelli AM, Schapira AH, Martinuzzi A, Carelli V, Rugolo M. Leber's hereditary optic neuropathy (LHON) pathogenic mutations induce mitochondrial-dependent apoptotic death in transmitochondrial cells incubated with galactose medium. J Biol Chem. 2003;278:4145-4150. 31. Zanna C, Ghelli A, Porcelli AM, Carelli V, Martinuzzi A, Rugolo M. Apoptotic cell death of cybrid cells bearing Leber's hereditary optic neuropathy mutations is caspase independent. Ann N Y Acad Sci. 2003;1010:213-217. 32. Wong A, Cortopassi G. mtDNA mutations confer cellular sensitivity to oxidant stress that is partially rescued by calcium depletion and cyclosporin A. Biochem Biophys Res Commun. 1997;239:139-145. 33. Kinnally KW, Peixoto PM, Ryu SY, Dejean LM. Is mPTP the gatekeeper for necrosis, apoptosis, or both? Biochim Biophys Acta. 2011;1813:616-622. 34. McCommis KS, Baines CP. The role of VDAC in cell death: friend or foe? Biochim Biophys Acta. 2012;1818:1444-1450. 35. Watson A, Lovelock JE, Margulis L. Methanogenesis, fires and the regulation of atmospheric oxygen. Biosystems. 1978;10:293-298. 36. Margulis L, Chapman M, Guerrero R, Hall J. The last eukaryotic common ancestor (LECA): acquisition of cytoskeletal motility from aerotolerant spirochetes in the Proterozoic Eon. Proc Natl Acad Sci U S A. 2006;103:13080-13085. 37. Vecino E, Hernández M, García M. Cell death in the developing vertebrate retina. Int J Dev Biol. 2004;48:965-974. 38. de Castro IP, Martins LM, Tufi R. Mitochondrial quality control and neurological disease: an emerging connection. Expert Rev Mol Med. 2010;12:e12. 39. Pan BX, Ross-Cisneros FN, Carelli V, Rue KS, Salomao SR, Moraes-Filho MN, Moraes MN, Berezovsky A, Belfort R Jr, Sadun AA. Mathematically modeling the involvement of axons in Leber's hereditary optic neuropathy. Invest Ophthalmol Vis Sci. 2012;53:7608-7617. 40. Klopstock T, Yu-Wai-Man P, Dimitriadis K, Rouleau J, Heck S, Bailie M, Atawan A, Chattopadhyay S, Schubert M, Garip A, Kernt M, Petraki D, Rummey C, Leinonen M, Metz G, Griffiths PG, Meier T, Chinnery PF. A randomized placebo-controlled trial of idebenone in Leber's hereditary optic neuropathy. Brain. 2011;134:2677-2686. 41. Carelli V, La Morgia C, Valentino ML, Rizzo G, Carbonelli M, De Negri AM, Sadun F, Carta A, Guerriero S, Simonelli F, Sadun AA, Aggarwal D, Liguori R, Avoni P, Baruzzi A, Zeviani M, Montagna P, Barboni P. Idebenone treatment in Leber's hereditary optic neuropathy. Brain. 2011;134(pt 9):e188. 42. Newman NJ. Treatment of Leber hereditary optic neuropathy. Brain. 2011;134:2447-2450. 43. Sabet-Peyman EJ, Khaderi KR, Sadun AA. Is Leber hereditary optic neuropathy treatable? Encouraging results with idebenone in both prospective and retrospective trials and an illustrative case. J Neuroophthalmol. 2012;32:54-57. 44. Sadun AA, Chicani CF, Ross-Cisneros FN, Barboni P, Thoolen M, Shrader WD, Kubis K, Carelli V, Miller G. Effect of EPI-743 on the clinical course of the mitochondrial disease Leber hereditary optic neuropathy. Arch Neurol. 2012;69:331-338. 45. Sadun AA, La Morgia C, Carelli V. Leber's hereditary optic neuropathy. Curr Treat Options Neurol. 2011;13:109-117. 46. Yu H, Koilkonda RD, Chou TH, Porciatti V, Ozdemir SS, Chiodo V, Boye SL, Boye SE, Hauswirth WW, Lewin AS, Guy J. Gene delivery to mitochondria by targeting modified adeno associated virus suppresses Leber's hereditary optic neuropathy in a mouse model. Proc Natl Acad Sci USA. 2012;109:E1238-E1247. 47. Lin CS, Sharpley MS, Fan W, Waymire KG, Sadun AA, Carelli V, Ross-Cisneros FN, Baciu P, Sung E, McManus MJ, Pan BX, Gil DW, Macgregor GR, Wallace DC. Mouse mtDNA mutant model of Leber hereditary optic neuropathy. Proc Natl Acad Sci USA. 2012;109:20065-20070. Sadun: J Neuro-Ophthalmol 2013; 33: 189-197 197 The Eleventh Annual Hoyt Lecture Copyright © North American Neuro-Ophthalmology Society. Unauthorized reproduction of this article is prohibited. |